DIRECT RELATIONSHIPS
Introduction
Whereas collateral relationships were described in great detail in the main monograph, direct relationships were treated more generally. The following section aims to redress the balance.
There are fewer kinds of direct relationship than collaterals and they are usually less complicated. This is because there are no half or first degree* forms, which are a common feature of all collaterals. Furthermore, all direct relationships are intergenerational while collaterals can be members of the same or different generations. Finally with direct relationships, gene transmission is forward and unidirectional, in contrast to collaterals whose connections run in both directions of time through common ancestors.
A further fundamental difference is that inbreeding plays a more active role in the formation of double and multiple direct relationships. Most double and multiple collaterals are possible without inbreeding, but with direct relationships all plural forms result from matings between fairly close relatives.
A puzzling feature of direct lines of inheritance is that they ostensibly appear to branch limitlessly in both directions. (Fig. 25). This is a gross oversimplification of the true situation. In reality, many of the branches into the past are only temporary because of inbreeding. This results in many ancestors being repeated several times in the pedigree causing their overall numbers to decline as we go back in time. Branches into the future also show a lack of permanence, with some coming to an abrupt end and others expanding more rapidly to compensate.
Fig. 25 also shows a further peculiarity of these relationships in that the subject (individual X♂) is directly related to all his ancestors' spouses (or partners) but not to those of his descendants. This rule partially breaks down when an ancestor later remarries and/or when inbreeding occurs among the descendants. Previous discussions on direct relationships can be found on here in the main monograph and here and here in this Supplement.
* Relationship increased by the presence of identical (monozygotic) twins.
Figure 25 Stylised Representation of Branching Direct Lines
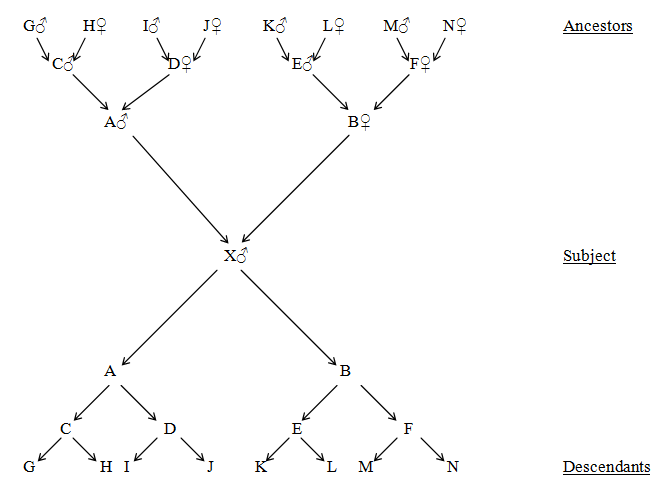
The simplest representation of direct relationship is a single line diagram (Fig. 26) which is theoretically continuous back to the earliest life forms through parent, grandparent and great grandparent etc. Immediately below the parent is the subject or proband (X) who is the designated person on whose behalf the lineage is being investigated. The extension forward in time from the proband consists of child, grandchild, great grandchild and so on. Any imaginary projection beyond the present into the future can never be guaranteed and the line may be cut short at any time. The termination of a line can be due either to the death of all the survivors of that line or their failure to have children. In the case of single sex lines, this can also happen when only members of the wrong sex are produced. Thus, lines of direct descent always have a definite past but an indefinite future.
In the following sections, coefficients of relationship (R) are given with each diagram and where close inbreeding is involved, F values are indicated and the R values are adjusted accordingly. The relevant individuals in each two-way relationship are highlighted in red.
Figure 26 General Representation of Direct Relationships

In addition to the specific types of ancestors and descendants shown above, direct relationships can also be classified in the following ways:
Further Categories of Direct Relationship
(a) Single direct relationships.
(b) Regular double and multiple direct relationships.*
(c) Irregular double and multiple direct relationships.*
(d) Irregular doubles combining direct and collateral relationships.
(e) Unilineal and bilineal direct relationships.
(f) Male, female and mixed direct lines of descent.*
* Extended (i.e. multigenerational) examples of (b), (c) and (f) from the British monarchy are given here, here and here respectively.
(a) Single Direct Relationships
The parent - child relationship is exclusively single, 'grandparent - grandchild' can be either single or double and the more distant direct relationships can be single, double or multiple.
Figure 27 Single Direct Relationships
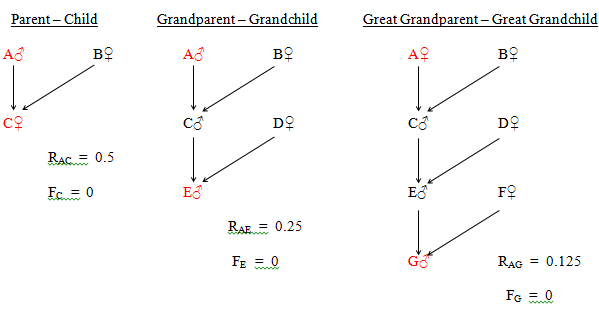
As stated earlier, direct relationships can only be double or multiple as a result of inbreeding. The double examples in Fig. 28 involve full sib and first cousin matings.
(b) Regular Double and Multiple Direct Relationships
Figure 28 Regular Double Direct Relationships

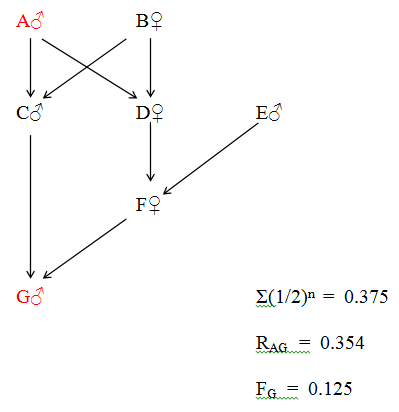
Figure 29 Regular Multiple Direct Relationship
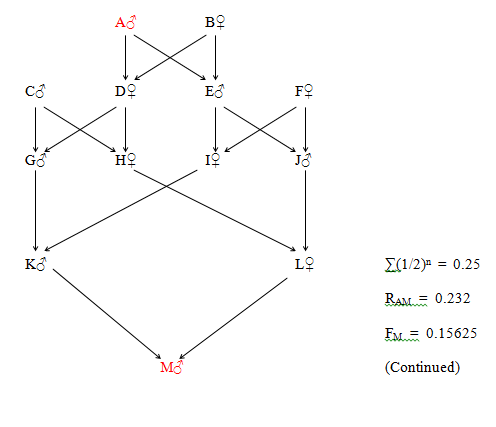
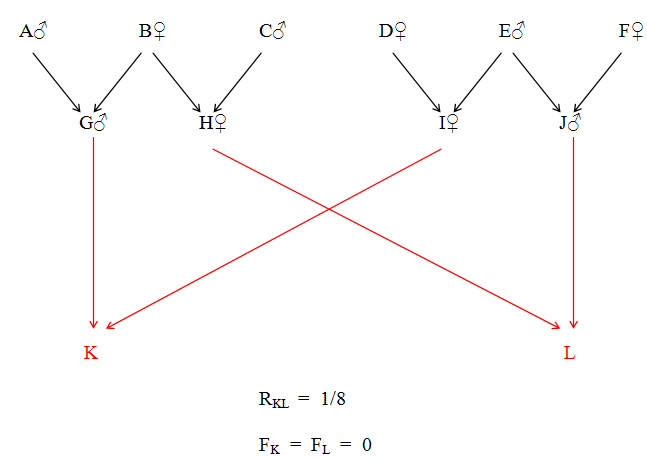
K and L are double first cousins and also double second cousins. The relationship of A to M is that of Quadruple Great Great Grandfather. The four paths between A and M are as follows:
A - D - G - K - M
A - D - H - L - M
A - E - I - K - M
A - E - J - L - M
In domestic animals and plants, inbreeding levels can be even more intense, leading to further links.
Figure 30 Octuple Great Great Grandfather (Three generations of full sib mating.)
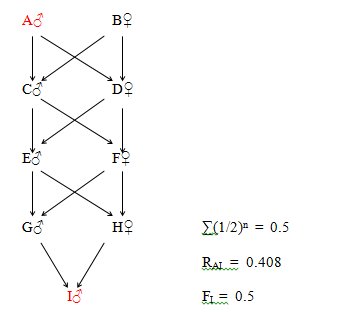
The eight paths between A and I are as follows:
A - C - E - G - IA - C - E - H - I
A - D - E - G - I A - D - E - H - I
A - C - F - G - I A - C - F - H - I
A - D - F - G - I A - D - F - H - I
Extended Regular Double Direct Relationships
Figure 31 John of Gaunt to Margaret Tudor (R = 0.0624)
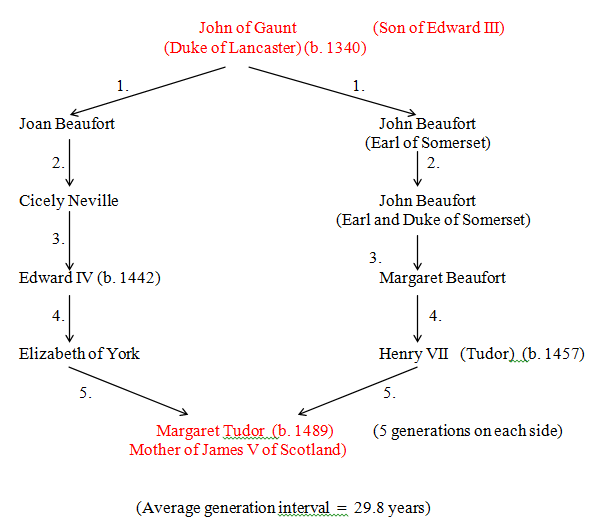
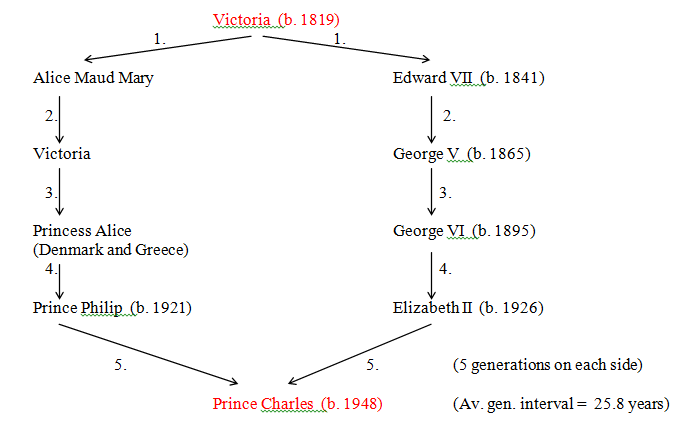
Figure 32 Queen Victoria to Prince Charles (R = 0.0624)
(c) Irregular Double and Multiple Direct Relationships
All the double and multiple relationships shown above have been regular. This means that the lengths of the various paths joining the ancestor and descendant are all equal. When the number of generations in one path gets out of phase with those in another, the double or multiple relationship becomes irregular, (i.e. asymmetrical). The significance of non-synchronised generations and variable generation intervals was covered in the main monograph here.
Irregular Double Direct Relationships
The first irregular double is the result of an uncle - niece marriage, which although illegal in Britain, has sometimes been allowed to happen.
Figure 33 Irregular Double Grandparent
The two unequal lines of descent are:
A - C - G (A is grandfather to G).
A - D - F - G (A is also great grandfather to G).
A more common irregular double results when a person marries his/her first cousin once removed. (Fig. 34). This is perfectly legal in the UK.
Figure 34 Irregular Double Great Grandparent
The two unequal lines of descent are:
A - D - G - K (A is great grandfather to K).
A - E - H - J - K (A is also great great grandfather to K).
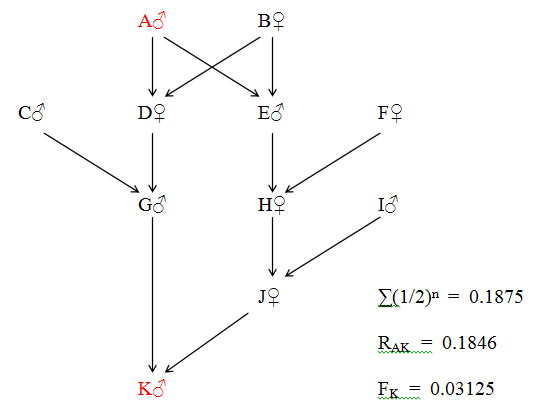
Figure 35 Irregular Multiple Direct Relationship
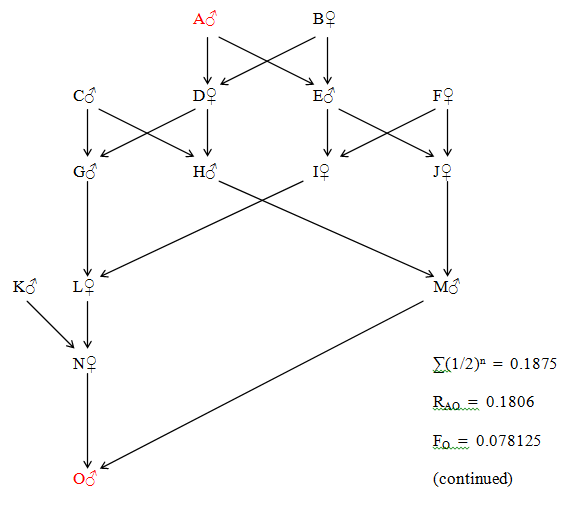
The four unequal lines of descent are:
A - D - G - L - N - O (5 generations) A - D - H - M - O (4 generations)
A - E - I - L - N - O (5 generations) A - E - J - M - O (4 generations)
Further irregular double direct relationships are shown later (Figs.56 and 57).
Extended Irregular Double and Multiple Direct Relationships
Generation intervals in different branches of the same family can get seriously out of step resulting in two individuals marrying who are several generations apart.
Extended Irregular Royal Doubles
Figure 36 Edward III to Richard (Duke of York) (R = 0.1556)
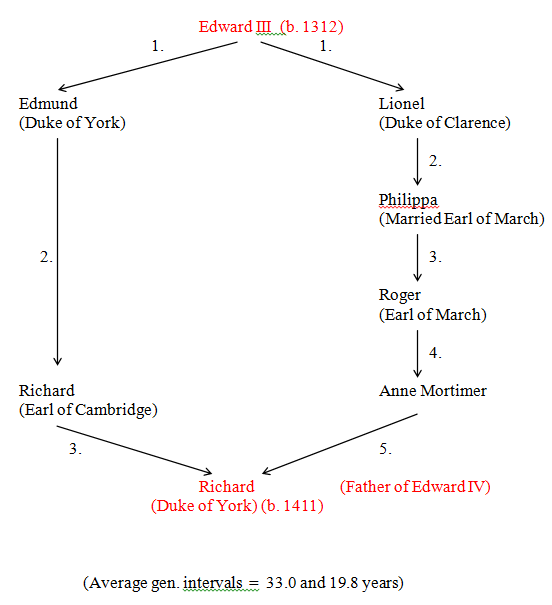
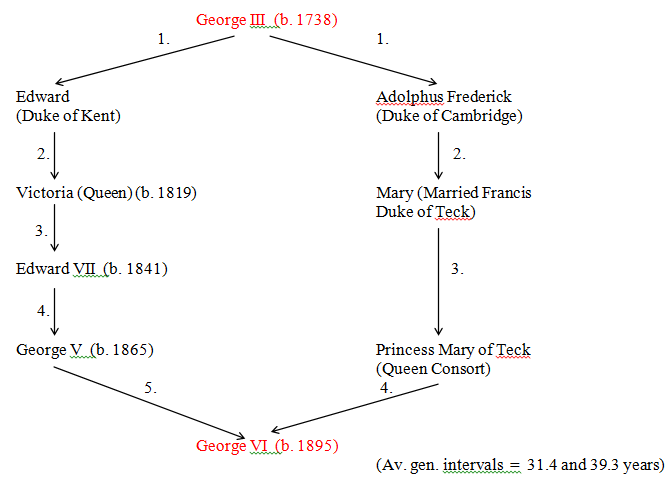
Figure 37 George III to George VI (R = 0.0934)
Extended Irregular Royal Multiple
There are several direct lines linking the Plantagenet kings with the Tudors and Stuarts. To simplify the diagram in Fig. 38, the following coding system has been used:
| A = | Edward III (Plantagenet) (b. 1312) | N = | Richard, Duke of York |
| B = | Philippa of Hainault | O = | Cicely Neville |
| C = | Lionel, Duke of Clarence | P = | John Beaufort (2), Earl and Duke of Somerset |
| D = | Edmund, Duke of York | Q = | Joan Beaufort (2) |
| E = | John of Gaunt, Duke of Lancaster | R = | Edward IV (House of York) (b. 1442) |
| F = | Katherine Swinford | S = | Margaret Beaufort |
| G = | Philippa, daughter of Lionel, Duke of Clarence | T = | James II (Stuart) King of Scotland (b. 1430) |
| H = | Roger, Earl of March | U = | Elizabeth of York |
| I = | Anne Mortimer | V = | Henry VII (Tudor) (b. 1457) |
| J = | Richard, Earl of Cambridge | W = | James III , King of Scotland (b. 1452) |
| K = | Joan Beaufort (1) | X = | Margaret Tudor |
| L = | John Beaufort (1), Earl of Somerset | Y = | James IV, King of Scotland (b. 1473) |
| M = | Margaret de Holand | Z = | James V, King of Scotland (b. 1512) |
Figure 38 Edward III to James V of Scotland (R = 0.0331)
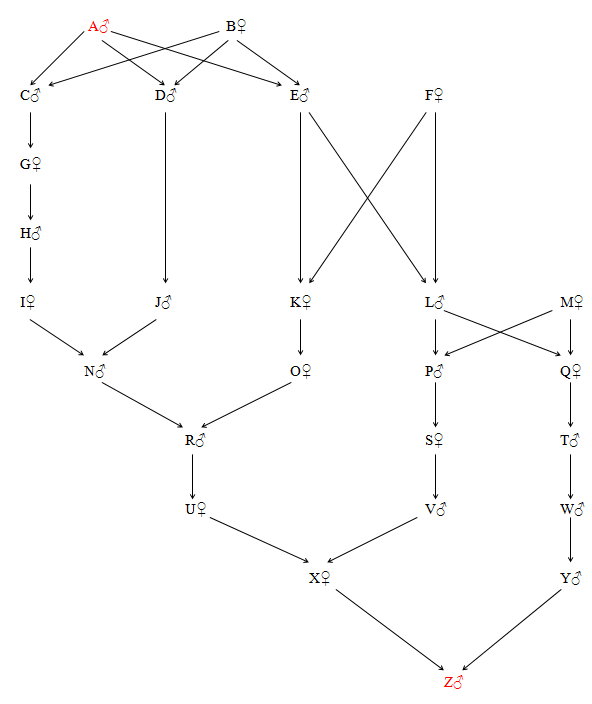
The five direct lines are as follows:
A - C - G - H - I - N - R - U - X - Z (9 generations)
| A - D - J - N - R - U - X - Z | - (7 generations each) | |
| A - E - K - O - R - U - X - Z | ||
| A - E - L - P - S - V - X - Z | ||
| A - E - L - Q - T - W - Y - Z | (FN = 1.56%) (FR = 1.95%) (FX = 0.51%) (FZ = 0.52%) |
(Average generation intervals = 22.2 and 28.6 years)
(d) Irregular Doubles Combining Direct and Collateral Relationships
These double relationships are unusual since the individuals are related in two different ways, directly and through earlier common ancestors. They are always associated with inbreeding.
Figure 39 Close Inbreeding (Incest)
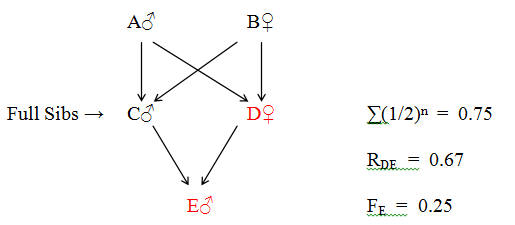
Direct relationship: D - E (Mother - Son)
| Collateral Relationship - | D - A - C - E | -(Aunt - Nephew) | ||
| D - B - C - E |
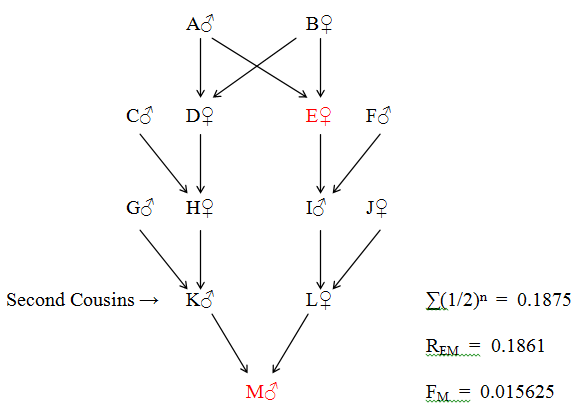
Direct Relationship: E - I - L - M (Great Grandmother - Great Grandson)
| Collateral Relationship - | E - A - D - H - K - M | - (Great Great Aunt - Great Great Nephew) | ||
| E - B - D - H - K - M |
There are further examples of mixed irregular doubles in Figs. 56 and 57
(e) Unilineal and Bilineal Direct Relationships
The genetic significance of unilineal and bilineal inheritance was explained in the main monograph here. Very briefly, any variance due to dominance interaction between alleles contributes to the phenotypic correlation between bilineal relatives but not between unilineals. Also the transmission of harmful genes between direct relatives is influenced by the type of lineal inheritance. In the case of direct relationships, the bilineal form is where the descendant is related through both of his/her parents to the ancestor and at the same time there is a certain probability that the ancestor is able to pass both members of a pair of genes at any locus down to his/her descendant by different independent routes.
Although all single direct relationships are unilineal, not every double direct relationship is bilineal. This is similar to the situation with collaterals (see monograph here and here). To be more precise:
(i) The parent - child relationship can only be single and unilineal (Fig. 41). There are no double or bilineal forms. All other single direct relationships are also exclusively unilineal (Fig. 41).
(ii) The double grandparent - grandchild relationship can only be bilineal (Fig.42). There are insufficient generations to allow unilineal doubles to form.
(iii) More distant, double (and multiple) direct relationships such as great grandparent - great grandchild and great great grandparent - great great grandchild can be either unilineal (Figs. 43 and 45) or bilineal (Figs. 44 and 46).
(iv) Since identical twins are exact copies of each other genetically, they must be regarded as a single individual when considering whether a direct relationship is unilineal or bilineal (Figs.43, 45 and 46).
(v) If an intermediate member of a multiple direct line is the sole transmitter of genes to later generations, he/she must do so by two independent routes for the original ancestor to be bilineally related to the final descendant (Fig. 46 - Types A and B). If he/she only passes genes through a single individual then the relationship is unilineal (Fig. 45 - Types D and E).
To simplify the following diagrams, most of the spouses or partners have been omitted and the relevant individuals and bilineal lines of descent are printed in red.
Figure 41 Single Direct Relationships (All Unilineal)
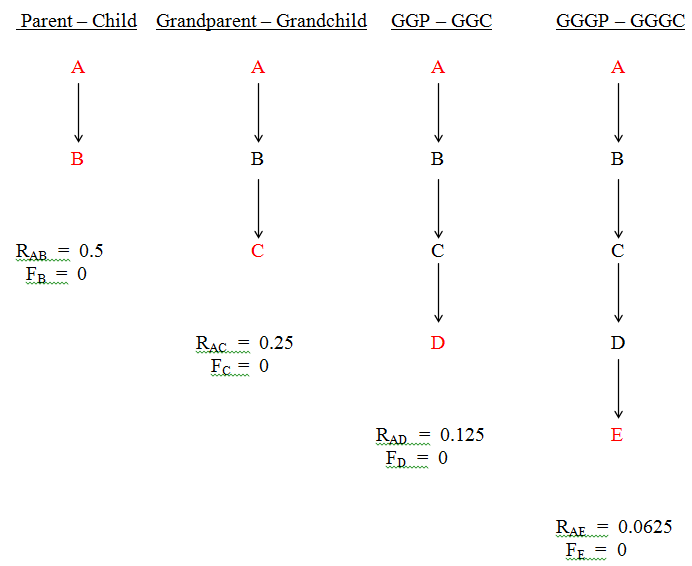
Figure 42 Double Grandparent - Grandchild (Bilineal Only)
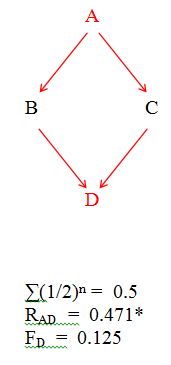
* B and C are only half sibs; if they had been full sibs then RAD would have been reduced to 0.447
Figure 43 Unilineal Double Direct Relationships
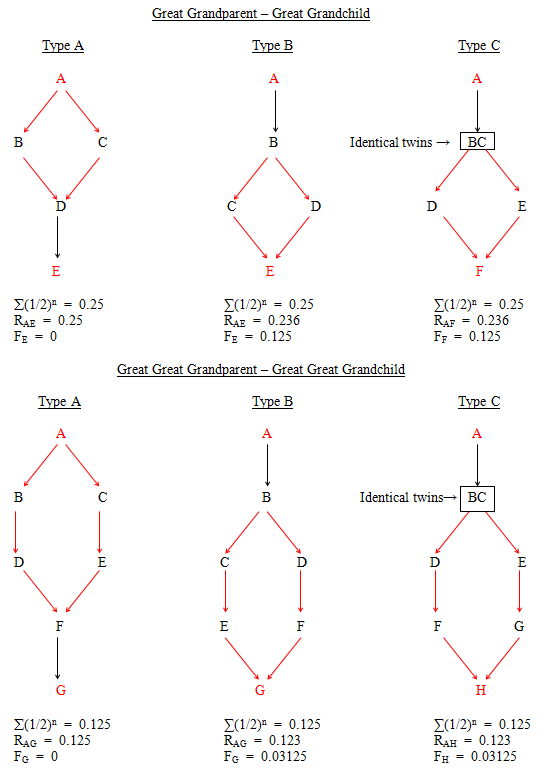
Figure 44 Further Bilineal Double Direct Relationships

Figure 45 Unilineal Multiple Direct Relationships
(Quadruple Great Great Great Grandpatents) **

Note: F in type A, G in type B and H in type C can also be replaced by identical twins.
Figure 46 Bilineal Multiple Direct Relationships
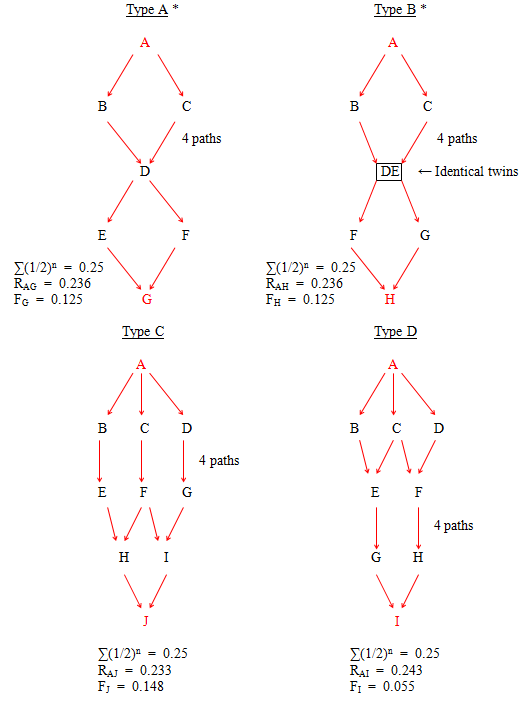
Note: F in type C and C in type D can also be replaced by identical twins without changing the outcome.
* Types A and B (Fig. 46) above both involve very close inbreeding and would normally only be found in animal breeding.
** Although there only appear to be two or three major lines of descent in each diagram in Figs. 45 and 46, there are in fact four in each case.
e.g. A to G above in Fig. 46 (Type A):
A - B - D - E - G
A - B - D - F - G
A - C - D - E - G
A - C - D - F - G
Also A to I in Fig. 46 (Type D):
A - B - E - G - I
A - C - E - G - I
A - C - F - H - I
A - D - F - H - I
The above topic (unilineal versus bilineal direct relationships) is mainly of academic interest since the parent-offspring relationship is the only direct relationship used in practice to estimate the heritability of traits. Estimates from more distant direct relatives take longer to realise and require greater allowances for non-contemporary environmental effects. Furthermore most of the double and bilineal direct relationships are complicated by the effects of inbreeding. It is more important in Mendelian genetics as shown earlier.
Not shown in the main monograph (except for a single example on here) is that some collateral doubles can also have 'variant' unilineal forms with the same degree of relationship. Most of these lack the symmetry of their bilineal counterparts. (For an example see Figs. 47 and 48).
Variant Double Collateral
Figure 47 Variant Form of Double Half First Cousins (Unilineal)

Figure 48 Conventional Double Half First Cousins (Bilineal)
f) All-Male, All-Female and Mixed Direct Lines of Descent
All-male direct lines of descent are characterised by the presence of the Y sex chromosome which is passed down from male to male virtually unchanged. This is known as patrilineal inheritance. Only gene mutations, which are very rare events, can cause changes to its genetic material. The reason for this is that for most of its length, the short Y chromosome has no homologous (i.e. equivalent) part on the much longer X chromosome. The X chromosome carries most of the sex-linked genes in mammals, the inheritance of which was described on here in the monograph. Since most of the male Y chromosome has no corresponding part on the X there can be no recombination of its genes by the mechanism of crossing over, which normally occurs with other chromosomes. Consequently, the Y chromosome remains unchanged over very long periods of time and can be used to track direct male lines right back into antiquity.
All-female direct lines have a similar feature which enables the same exercise to be carried out. The mitochondria, which are the small organelles in the cytoplasm of cells, responsible for energy production, are not controlled by the genes in the cell nucleus. They have their own genetic determinants which are passed down through the generations unchanged in the cytoplasm of the female egg cells. Both male and female offspring receive their mitochondria from the mother's ovum. Males can not pass mitochondria through their sperm cells so mitochondria remain a distinguishing characteristic of the female line. (Matrilineal inheritance).
The following diagrams (Fig. 49) illustrate five different kinds of direct line. In no. 1, the Y chromosome is passed down through the male line unchanged from A to H. In no. 2, the mitochondria are passed down through the female line in the same way. In no. 3, H male receives his mitochondria unchanged from A female but the transmission goes no further. In no. 4, H female does not receive a Y chromosome, but can trace her male line ancestry if she has access to her father's (or her brother's, paternal uncle's or paternal uncle's son's) Y chromosome. Finally, no. 5 line, where males and females alternate, can supply no useful information about distant ancestors.
Although only eight generations are shown in the diagrams, the patterns can continue indefinitely.
Extended lines 1 to 4 enable the identification of common ancestors of large numbers of people in the distant past. As a result, it is possible to trace both the origins and migrations of different ethnic groups over long periods of time. Individuals who are highlighted in red have the same type of Y chromosome and those in green share the same mitochondria.
Figure 49 Male, Female and Mixed Direct Lines
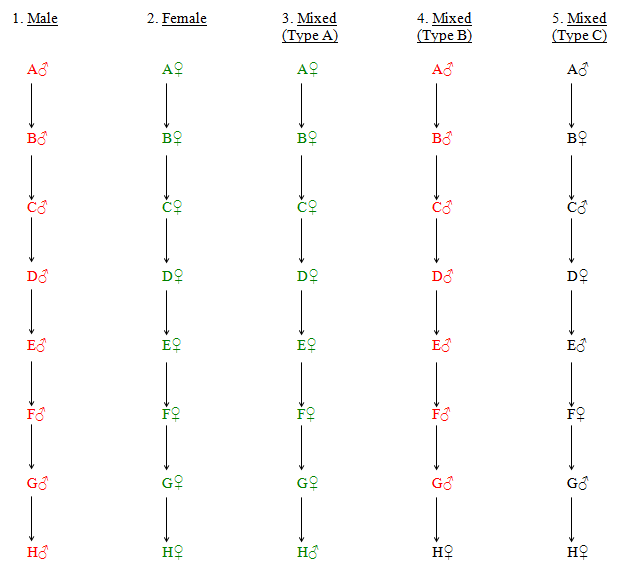
Providing the rule of male primogeniture is observed for hereditary titles, and family surnames always follow those of the male parents, then specific titles and family names for males will tend to be linked with particular Y chromosomes.
The survival of a direct line of the British monarchy for over 1200 years and 40 generations has only been possible because, in the absence of a male heir, female sovereigns and non-sovereign female intermediates have been accepted. In spite of this there are five separate runs of British kings, each presumably with its own distinctive Y chromosome. The occasional substitution of the king's first-born son with other males in the direct line, such as brothers and brothers' sons, does not affect the Y chromosome link. However, even though the British monarchy is not exclusively male, Prince Charles can theoretically trace at least one male line right back into antiquity, but it may not be completely royal.
Prolonged mitochondrial lines do not fare very well in royal families. Recently, the Dutch royal family has had a direct succession of three female sovereigns: Wilhelmina, Juliana and Beatrix. Strangely, although the Queen and Prince Philip both have the same relationship to Queen Victoria, only Philip inherited her mitochondria. (See Fig.32).
The total number of sovereigns of England to James I and of the United Kingdom thereafter, in the direct line from Egbert to Elizabeth II, is only 25. The total of all the sovereigns during that period, is 62 (including Lady Jane Grey, and Mary II who reigned with her husband William III). Apart from the usurpers (King Cnut, his two sons and Harold II) and the directly-related Norman kings (William I and Henry I), the remainder (31), exactly one half of the total, are only colaterally related to Elizabeth II. In contrast, most (54) of the 'true' (i.e. blood related) English and British sovereigns, with the exception of the early Norman kings (William I and II, Henry I and Stephen), have a direct relationship with King Egbert; because they are all his descendants. The theme of tracking a direct line through the ages was used in John Buchan's (1921) book, ' The Path of the King', which follows a hypothetical succession from a disinherited Viking prince down to Abraham Lincoln. However, in this case the symbolic link was not a Y chromosome but an ancient gold ring.
Figs. 50 to 54 show examples of long unbroken sequences of British kings. Kings' names are printed in red and those of non-sovereign intermediate males in black. The direct lineage and average generation interval of each uninterrupted male section of the royal line are also indicated in red.
Figure 50 The Saxons (8 generations - 16 kings)

* The direct royal line was passed on through Edmund II's granddaughter, Margaret, Queen (consort) of Scotland. Margaret's great grandson was Henry II (Plantagenet).
Figure 51 The Plantagenets (11 generations - 14 kings)
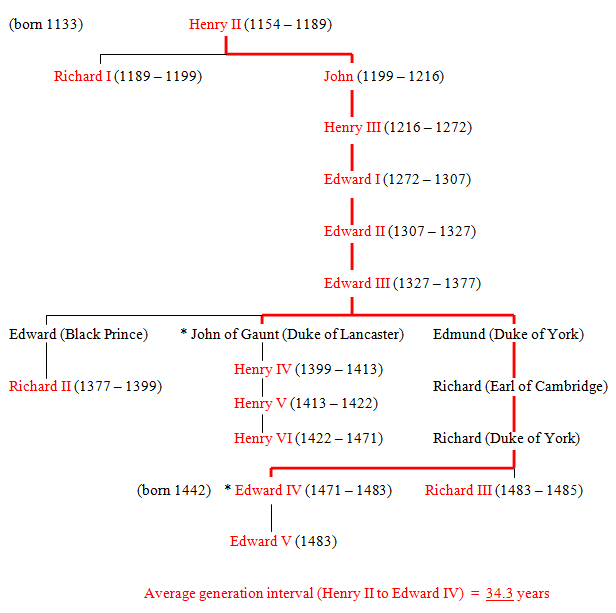
* The succession from hereon is quite complicated. The direct line was carried on through three separate strands involving the females: 1) Elizabeth of York, Edward IV's daughter; 2) Joan Beaufort, John of Gaunt's granddaughter and 3) Margaret Beaufort, John of Gaunt's great granddaughter. Eventually, these three independent links all came together in the person of James V (Stuart) King of Scotland. (See Fig.38 and footnote to Fig 52).
The direct line through Margaret Beaufort also included Henry VII, the only Tudor in the direct line. John of Gaunt's part in the royal succession was not through Henry IV but through Edward IV, Henry VII and the Scottish Stuarts. (See Fig. 38). Excluding the purely Scottish kings (James II, III and IV), Edward IV and Henry VII are the only two English/British kings (in the direct succession from Egbert to Elizabeth II) that occur in two separate parallel lines. (See Fig. 31). The rest all form a linear series.
Figure 52 The Scottish Stuarts (7 generations - 7 kings)

* The direct lines for both the Scottish and English crowns were passed on through James V's daughter, Mary Queen of Scots, who was heir to both thrones. This resulted in her son, James VI, becoming King of both countries in 1603. James VI of Scotland and I of the United Kingdom passed on the crown directly to George I (Hanover) through two female intermediates, Elizabeth and Sophie (his daughter and granddaughter respectively). Thus, James I (Stuart) shares the distinction with Henry VII (Tudor) in being the only member of his dynasty to be included in the direct royal line of England and the UK. All the earlier Stuarts were just sovereigns of Scotland and the later ones were not in the direct line.
Figure 53 House of Hanover (5 generations - 5 kings)

The direct line was passed on through Edward, Duke of Kent's daughter, Queen Victoria to her son, Edward VII.
Figure 54 House of Saxe-Coburg-Gotha/Windsor (3 generations - 4 kings)

George VI passed on the crown to his daughter Elizabeth II. If Prince Charles, Prince William and Prince George inherit the crown, their Y chromosomes will have come from the Mountbatten dynasty.
Y Chromosome Inheritance
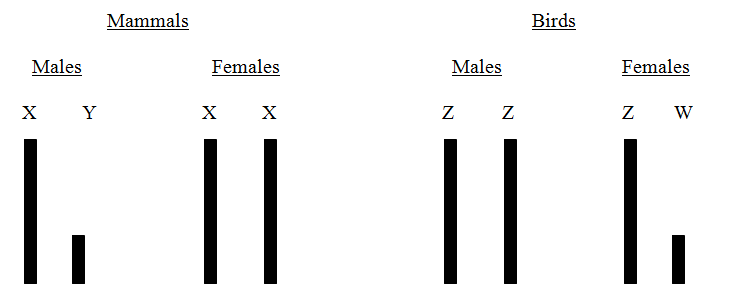
Unlike the X chromosome, the inheritance of genes on the Y chromosome in humans was not covered in the main monograph. Genes on that part of the Y chromosome, which has no equivalent section on the X, can only code for proteins and characters that appear exclusively in males. The most important of these is SRY (sex-determining region Y) which triggers the development of male characters in the embryo. The rest are fewer in number and perhaps less interesting than those on the X. However, in birds, where a similar situation exists with regard to sex-linked inheritance, a fascinating example of 'Y chromosome' inheritance has been found.
In birds, the smaller sex chromosome is called W and the larger one Z. Furthermore, the sexes are reversed in the morphology of their sex chromosomes.
Figure 55 Diagrammatic Representation of Sex Chromosomes in Birds and Mammals
The Common Cuckoo (Cuculus canorus) is a brood parasite of other small birds such as Dunnocks, Meadow Pipits and Reed Warblers. The female cuckoos are divided into several distinct groups or gentes. Each group chooses the nest of one particular host species and, apart from the Dunnock, lays an egg with a similar shell colour to the host species. When the young cuckoo hatches it pushes the chicks and unhatched eggs of the host species out of the nest and is reared by the foster parents. However, the male cuckoos are not divided in this way and seem to be able to mate with any group of females without affecting the shell colour or nesting habits of their female offspring. It has been suggested (Dawkins, 1998) that the genetic explanation of this phenomenon is that the genes for nest preference and egg shell colour are located on the short W chromosome. Since the male only has Z chromosomes he can not influence these characteristics. The Dunnock, has probably only recently been parasitised and has not yet learned to recognise the different coloured eggs of the cuckoo.
Thus, conditions or characters that appear only in males in mammals and only in females in birds, may be determined by genes on the Y or W chromosomes. However, such conditions should not be confused with other sex-limited characters, such as milk yield or semen quality in mammals, which are controlled by sex hormones and whose genetic determinants are present in both sexes.
Matings Between Directly-Related Individuals
Marriage between living direct relatives is forbidden and most forms are classed as incestuous. Parent - child and grandfather - granddaughter marriages are both illegal and incestuous. Grandmother - grandson marriage is illegal but not on the list of incestuous relationships. Great grandfather - great granddaughter marriages are listed as illegal in Scotland but not in England. However, there is a tradition of 'implied generic inclusion'. For example, in the early forbidden lists, although half brother - half sister marriages were not included, the words brother and sister were generally understood to include all types of siblings. Similarly, grandfather and grandmother are general terms that could also include earlier ancestors.
With modern technology, the long-term preservation of viable sperm and eggs is now possible; giving a wider span of generations for potential parents. More precise legislation may be necessary in the future to clarify the legal situation regarding matings between distant direct relatives. The relationship with ancestors earlier than great grandparents is less than that between first cousins, but wide cross-generation breeding could cause some weird legal and relationship complications.
Even though sexual relations between close direct relatives are classed as incestuous, with severe penalties, they still happen quite frequently with harmful results. Any children from these liaisons have some very strange irregular double relationships with their predecessors. (See Figs. 56 and 57) .
Figure 56 Father - Daughter Incest
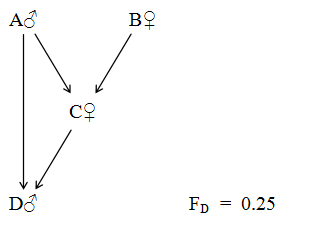
Relationships to Proband D
A♂ = Father and Grandfather ∑(1/2)n = 0.75 RAD = 0.67
C♀ = Mother and Half Sister ∑(1/2)n = 0.75 RCD = 0.67
Figure 57 Grandfather - Granddaughter Incest
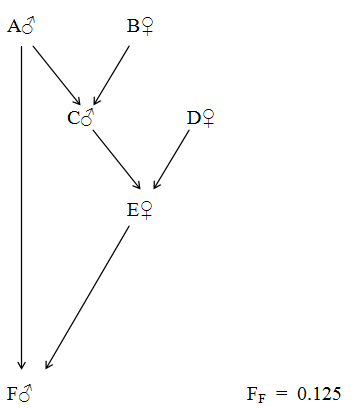
Relationships to Proband F
| A♂ | = | Father and Great Grandfather | ∑(1/2)n | = | 0.625 | RAF | = | 0.59 |
| C♂ | = | Grandfather and Half Brother | ∑(1/2)n | = | 0.5 | RCF | = | 0.47 |
| E♀ | = | Mother and Half Niece * | ∑(1/2)n | = | 0.625 | REF | = | 0.59 |
* It is not uncommon for nephews and nieces to be older than their uncles or aunts. It happens when a woman has further children of her own after her grandchildren are born. (See here). However, in this case the mother is half niece to her own son!